Practical Genetics, Step 2 --
The practical breeder's second job is to make a reasonable guess about the mode of inheritance for the trait.
There are, basically, four modes of inheritance you may see for a given genetic trait: simple dominant, simple (but either autosomal or sex-linked) recessive, polygenic, or other (ie, two- or three-gene traits or incomplete dominance or something fairly peculiar, such as trinucleotide-repeat traits).
The inheritance pattern has not yet been determined for many (most) of the genetic problems seen in dogs, particularly those found at very low frequencies. Therefore the field is wide open! You may be able to serve your breed (and your own kennel) well by establishing the mode of inheritance for some problem that occurs in your breed. This is very nice, if possible.
While we're at it, let's not forget that there are also non-disease traits with which you will certainly need to work while trying to achieve your breeding goals. Many or most "type" traits, such as correct ears, are certainly polygenic -- ear shape, size, set, and amount of furnishing are all involved in creating the correct overall type for the ears alone, for example, before we even get to the rest of the head -- and these type traits will be handled like any other polygenic trait. Established "top" breeders know exactly what they're doing when it comes to handling polygenic "type" traits -- though polygenic traits like hip dysplasia are much harder, because they don't show on the outside, they are actually handled the same way: by looking at which animals produce what; by looking at close relatives; by looking at sibs as well as grandparents (breadth in the pedigree as well as depth); and so forth.
Some type characteristics, such as coat texture, whether a nose is "butterfly", occurrence and extent of ticking, or the presence of extra dew claws, are probably relatively simple in inheritance. In general, if you could graph type most easily on a normal (bell) curve, then it's likely to be polygenic; whereas if you can identify animals as either Yes or No for a trait, it's likely to be simple. (These kinds of traits might also be referred to as quantitative vs. qualitative.)
There are two basic ways to get good information about the mode of inheritance of a relatively simple trait or disease.
a) Pedigree analysis.
b) Test-matings.
If somebody else has done the work for you, great! But too often a source will list a trait as "a simple recessive" on the basis of one family of dogs, or even on just one litter of puppies, and then you really don't know whether the conclusion was sound. A lot of the time, a source gives you no information at all on what data led to a particular conclusion, and it is not safe to assume the conclusion was sound. It is appalling how often an important conclusion is based on bad data. It's worth doing a literature search first thing when you get interested in a trait, but always, always, always be sure either to find support from a really trustworthy source, or to track down the actual data yourself before you just accept somebody else's conclusion. Hutt, Willis, and the lit cited sections for some of the Ruvinsky & Sampson articles are places to go if you're trying to track down information, for example, and I consider Hutt the most trustworthy in his conclusions about inheritance patterns. The internet can be helpful, but often you just get conclusions passed along from site to site with no way to tell whether the conclusions are based on good data. A general paucity of information about lots of traits means that often a breeder will have to generate or collect and then analyze data herself if she wants to have good information available.
I) Pedigree Analysis:
To determine mode of inheritance for a particular trait via pedigree analysis, what you will need when working with phenotypic traits is a really thorough pedigree for your dogs. This is a pedigree that should include the full sibs of each dog in your breeding program, and the full sibs of the parents of each such dog, and the full sibs (preferably) of the grandparents of each such dog. It is nice to include half-sibs also, if possible. Pedigrees of this sort allow you to figure out what mode of inheritance can reasonably be predicted to underlie the phenotypic traits you see in your dogs.
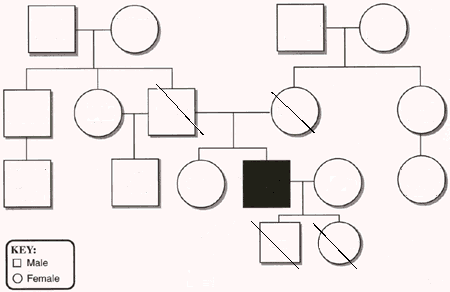
One way to show this sort of expanded pedigree, once you have it, is demonstrated by Padgett (1998). It looks something like this:
Only usually a lot more extensive. Padgett's sample family trees are huge and contain a lot more information. Here, the trait in question is most likely a simple recessive: the black square is affected, strike-throughs would be obligate carriers, and everybody else shows normal phenotype and has unknown genotype (they could be carriers -- indeed, some of them must be carriers). Add enough animals to this family tree and you gradually get a heck of a lot of information in one place.
Once you have an expanded pedigree in front of you, you can at the very least make an educated guess about the inheritance pattern for this disease – or conceivably you may decide the problem is probably not genetic and start looking for a causative environmental factor.
Check first for the simple inheritance patterns: look for a simple autosomal recessive pattern. Then autosomal dominant. Sex-linked recessive. Sex-linked dominant. Incomplete dominant. Consider the possibility of incomplete penetrance. Consider the (rather likely) possibility that the problem is polygenic.
Here’s what you should see in a family tree if a disease or trait turns out to be a straight-up no-frills autosomal dominant:
a) Affected dogs always have one, but probably only one, affected parent.
b) The line of affected dogs goes straight back, skipping no generations, to the dog that suffered the original mutation. Lack of penetrance will create "holes" in this line, but if penetrance is very weak, then you haven't got a simple dominant anyway.
c) Mating an affected bitch to an affected dog should NOT result in all affected offspring. (Roughly 25% should be unaffected.) Small litter sizes will obviously mess up expected Mendelian ratios. You can flip a coin four times and get heads four times in a row, but that doesn’t mean you couldn’t have gotten tails.
c) If the disease is a sex-linked dominant, all daughters of an affected male will be affected, but none of the sons. This probably won’t be the case, but it’s theoretically possible that you’ll find a real no foolin’ sex-linked dominant.
d) Roughly half of the offspring of an affected dog will be affected if crossed to a normal dog, and the other half will be normal. Again, with the small litter sizes of the Cavalier, you’re not likely to be able to assess this expectation with fewer than two or three litters.
Here’s what you should see in a family tree if a disease turns out to be a simple autosomal recessive:
a) Affected dogs usually have normal parents, especially if the trait is early-onset and actively
being selected against by breeders.
b) The trait is clearly familial, but skipped generations are common.
c) Roughly ¼ of the puppies produced by two carriers will be affected
d) A carrier mated to an affected dog will produce approximately half affected puppies.
e) Two affected animals mated together will produce 100% affected puppies.
Here’s what you should see in a family tree if a disease turns out to be a sex-linked recessive:
a) Affected dogs usually have normal parents, especially if the trait is early-onset and actively
being selected against by breeders.
b) The trait is clearly familial, but tends to skip from great-uncle to great-nephew.
c) More male animals than females are affected.
d) Roughly ¼ of the puppies produced by a carrier female bred to a normal male will be affected, but they will all be male.
e) All of the daughters produced by an affected male will be carriers.
f) Two affected animals mated together will produce 100% affected offspring.
Here’s what you should see in a pedigree if the problem you are considering is polygenic:
a) Affected dogs usually have normal (or much less severely affected) parents, if
the trait is being selected against by breeders.
b) The risk of getting affected puppies rises with increasing severity
of parental symptoms. For example, the worse the hips of the parents, the more
likely some degree of dysplasia will appear in the puppies.
c) The risk of getting affected puppies rises as the number of affected siblings
already produced increases. That is, the more of the older sibs are
affected, the greater the risk that younger full-sibs will also be affected. This is not the
case for a simple recessive, where the risk in a carrier-carrier breeding is always 25%
for each puppy produced, regardless of the number of affected sibs already produced.
d) The risk of getting affected puppies declines sharply as the relationship to the
affected individual(s) becomes more distant. That is, the risk of producing
affected puppies if the sire is affected is much higher than if the the sire was clear but
the grandsire was affected; and a whole lot higher than if only the great-grandsire was
affected. This is not the case with a simple recessive, where risk does not get "diluted"
in the same way -- if the disease-causing allele is present, then it is present, period.
e) The sex less often affected is more likely to pass on the trait if it is affected.
For example, if more males than females develop MVD, which is the case in Cavaliers,
then if you have a female who does develop the problem, she is probably a worse
breeding risk than an affected male would be. This is because it takes, theoretically,
a higher genetic "load" of deleterious alleles for her to be affected in the first place -- and
she can pass on those alleles.
As a rule of thumb, metabolic defects – such as tyrosinase deficiency in the Chow – are probably controlled by some simple single-gene mechanism. When you have no information about a trait, then you may want to guess that a trait is simple, or relatively simple, in inheritance if it seems to be a metabolic disorder. Any disease caused by the dysfunction of a specific enzyme is one I would call metabolic.
Alternatively, complicated anatomical defects – hip and elbow dysplasia, bloat, and so forth – are probably going to prove to be polygenic traits. So are many cancers. This is particularly true if environmental factors play a substantial role in determining the occurrence of the trait, as with the joint dysplasias (diet, exercise) and, for example, bloat (temperament, whether the food bowl is raised). This is one reason I doubt, despite guesses to the contrary (http://www.canine-genetics.com/pedigree.htm), that bloat is controlled by any simple single-gene mechanism. It is not true that Occam's razor indicates that the simplest possible explanation must be preferred, particularly when so many characteristics of a trait suggest polygeny over simple genetic causation. If you must postulate incomplete penetrance to force your data to fit a single-gene model, it's time to either get more data and so increase your sample size, or look for alternate hypotheses -- usually polygeny or other complexity. Or preferably both.
Some apparently-anatomical traits really do have a simple metabolic origin, such as Malamute dwarfism, for example, and probably all the other forms of dwarfism as well. You aren't shortening every bone independently to create a dwarf, but interfering with a single hormone that in turn affects all bone growth. So this trait, which might first strike a breeder as complex, is actually simple. The categories aren’t clear cut. That’s one reason why I suggest them as rules of thumb, not set-in-stone laws of nature.
There are other modes of inheritance than those expected from fully simple vs. truly polygenic.
Here's what a pedigree should look like if a trait is controlled by a two-gene system in which both genes must be recessive homozygous for the trait to manifest itself (only aabb is affected, not AaBb or Aabb or AABB or whatever):
a) Affected puppies should have normal parents.
b) Carrier - to - carrier breedings should yield 1/4 to 1/16 affected puppies. Of
course, you would need a pretty large sample size to assess whether you were getting
roughly these ratios, but you should get substantially fewer affected puppies than the
constant 1/4 you'd expect from a single-gene system.
c) The trait is clearly familial, but quite rare in the affected lines -- unless the trait
is actually common in the breed (i.e., there is a huge number of carriers).
A few canine diseases appear to be inherited in two- or three-gene systems, including one form of epilepsy in Springer Spaniels and Standard Schnauzers. In the case of the epilepsy, one of the genes involved also appears to be sex-linked, or possibly part of the mitochondrial DNA. More diseases will probably be found to follow a two- or three-gene pattern in the future.
Here's what you might see in a family if a problem is caused by a trinucleotide repeat disorder:
a) The first recorded instance of the disease in a family may report relatively mild symptoms
and / or a late age of onset.
b) The severity of the symptoms, or the age of onset, or both, get worse as
generations pass.
c) The "penetrance" of the trait seems to increase as generations pass.
d) The disease might appear to be inherited in a "recessive" manner at first and then
shift, as generations pass, to an apparently more "dominant" mode of inheritance.
e) The disease itself might appear to be passed down from one sex, although the potential
to be a "carrier" (the premutation) might appear to be passed by the other sex.
There are no trinucleotide repeat disorders yet known for dogs, as far as I know. Since there are some known for humans, the potential for this kind of disorder to occur obviously exists. All trinucleotide repeat disorders known in humans -- I think -- involve neurological syndromes.
II) Test-Mating
To determine the mode of inheritance of a particular trait by test-mating, you must be willing to deliberately produce affected puppies. Whether you will be willing to do this depends on how horrible the trait is, how you feel about culling affected puppies, and most of all how important you think it is to determine the mode of inheritance of the trait.
Remember that if you go along breeding dogs without knowing the mode of inheritance, you are risking the production of affected puppies with each litter you produce and so is everybody else, forever. You will never, for any litter, have a really good idea how likely you are to produce affected puppies. In contrast, if you can once determine the mode of inheritance of the trait, you should be able to drastically reduce the risk of producing affected puppies for everyone, including yourself. If the problem turns out to be a simple single-gene trait, you have also set the stage for a possible attempt to develop a genetic test for the disease. Once a genetic test exists, it will never be necessary to produce another affected puppy again.
The best book I know of to read to get a really good look at the kind of data you need from a test cross, by the way, is Hutt's Genetics for Dog Breeders.
Now, here's what you need to do if you decide to do a test cross: keep and breed at least one affected animal. There are ways to "cheat" if it's impossible to raise an affected puppy to breeding age (you can use carrier-to-carrier breedings) but it's usually much more straightforward to use affected animals if possible.
Suppose you have a dog who is born with, oh, microphthalmia. This is listed by Padgett as a defect that is present at birth (ideal) and of unknown mode of inheritance. It is a very simple type of defect (small eyes), which strongly suggests that it could be controlled by a single gene (ideal), and it does not kill the animal (vital, if you're going to raise and breed it). You can also be fairly confident going in that this is not a dominant trait -- not unless it is so variable in expressivity as to be invisible in the presumably normal parents. Probably this is either a single-gene recessive or a polygenic trait and it seems like a good candidate for a single-gene trait.
Raise the puppy. When he is old enough, breed him to his mother or to any other bitch known to have produced microphthalmia (any obligate carrier will do, but if his mother is the only proven carrier to which you have access, she's the bitch you're going to have to use). If this is a recessive trait you know precisely what you should get in the puppies: half normal and half affected. If this is a Cavalier litter, you won't get enough puppies from one litter to reliably assess ratios. It will be best to breed this dog back to his mother several times, or to several obligate carriers at the same time, to increase your sample size. If you're breeding Irish wolfhounds and expect 14 pups per litter, it may not take more than one or two litters to get the job done, you lucky guy.
You can also breed this dog to any full sisters. If they are carriers, again you should get a fifty : fifty ratio of normal to affected puppies. If they are clear, you won't get any affected puppies (and if you have produced enough puppies, you may be able to establish that the sisters are clear, which means you can use them in your breeding program quite safely, perhaps replacing their carrier mother.) Padgett lays out table after table of probabilities, from which we can easily get the information that if a sister produces only normal puppies to her affected brother, then she has a nearly 94% probability of being clear if she produces four puppies and a nearly 97% chance of being clear if she produces five puppies.
If you cross this dog to his mother or a sister and get both affected and normal puppies, then all the normal puppies should be obligate carriers, if this is a simple recessive. To cinch the deal, cross two of these carriers. What you should get is 25% affected puppies and 75% normal. Do several crosses of this kind if you can, so as to boost sample size.
If you take two affected animals -- now that you have several -- and breed them together, then you should get 100% affected puppies. Try it for two litters -- it's virtually conclusive and puts the finishing touches on your hypothesis of autosomal recessive inheritance.
Are these patterns what you see? Remember that expected Mendalian ratios will almost never be met exactly in real life. If you get these results:
Affected dog x normal mother = 2 affected puppies + 3 normal puppies
Affected dog x normal full sister = 3 affected puppies + 1 normal puppy
Affected dog x normal full sister = 2 affected puppy + 1 normal puppy
then what are you to conclude? You should have gotten 1/2 affected puppies for all these breedings. You didn't. What does this mean?
Let's assume that both sexes are affected at pretty even rates, so that you decide that this is looking like an autosomal trait rather than a sex-linked trait. You can lump all these litters together because they are all the same kind of breeding (affected to carrier). What you have is a 7 affected : 5 normal ration, which is not quite fifty : fifty. Is it close enough to be compatible with Mendelian expectations? You can find out by using a very simple chi-square test. The demonstration doesn't use the same example, but of course it uses the same method you would use on the data above.
Then you can also do a chi-square test for carrier x carrier breedings to see if you got a 25% affected : 75% normal ratio.
Finally, you can make sure that when two affected dogs are bred together, that all the puppies are affected.
If you find that your results fit Mendelian expectations, great! You have established a strong probability that this trait is a simple autosomal recessive. Assuming all three types of crosses (affected x carrier, carrier x carrier, affected x affected) fit expectations, you can be quite confident that you are correct in this conclusion. Everyone in your breed will benefit from your work -- congratulations!
If you find that your results do not fit Mendelian expectations, it's time to ponder what might be going on.
In the example above, you see "too many" affected puppies to really fit the fifty : fifty expectation. However, if you ignored litters to normal full sisters in which no affected puppies occurred, then you would expect to under-count normal puppies. Suppose that this litter was also produced:
Affected dog x normal full sister = 0 affected puppies + 3 normal puppies.
This sister might be clear (and unable to produce affected puppies) or she might be a carrier and she just "flipped heads" three times, which is quite possible. You can't justify lumping this litter in with the other three litters because you don't know that it was an affected x carrier breeding. That is why it would be best to breed this dog back to his carrier sisters once more -- that would increase your sample size and you would know for sure you were doing affected x carrier breedings. However, you can also apply a correction factor to compensate for undercounted normal puppies. The method for this is also described here.
But suppose the ratios you got were really far off expectation. What then? For example, suppose you got this result:
Affected dog x mother = 4 affected puppies + 1 normal puppy
Affected dog x mother (second litter) = 3 affected puppies + 0 normal puppies
Affected dog x normal full sister = 2 affected puppies + 2 normal puppies
Affected dog x normal full sister = 3 affected puppies + 1 normal puppies
Now you have a ratio of 12 affected : 4 normal. This is way off the expected fifty : fifty ration. What could be going on? You're producing "too many" affected puppies. Could this be a dominant with incomplete penetrance? Is there perhaps some eye characteristic which "normal" dogs show which might represent a minimal expression of microphthalmia? Try an affected x affected breeding -- try several. If this is really a dominant, you should get 25% normal puppies -- or somewhat more, given that this trait seems to lack complete penetrance, if it is a dominant. Or could this just a sample-size problem? 16 puppies isn't very many -- you may have to grit your teeth and do a few more breedings.
Or suppose you got these results:
Affected dog x mother = 1 affected puppy + 5 normal puppies
Affected dog x normal full sister = 1 affected puppy + 4 normal puppies
Affected dog x normal full sister = 4 affected puppy + 4 normal puppies
Now you have a ratio of 6 affected : 12 normal. This is not enough affected puppies to meet expectations for a single-gene autosomal recessive. What could be causing this deviation? Other than low sample sizes? This is when you might consider a two- or three- gene system. It's not a lot harder to build and work with possible two-gene models. For example, suppose this was a two-gene recessive system, like this:
Dogs with genotype aabb are affected. Dogs with any other genotype are unaffected.
Then if you do an aabb x AaBb cross, you should get 1/4 affected puppies. It's this kind of cross which might be lowering the number of affected puppies you're getting from affected x carrier crosses. Here's how the Punnett square would look, if you want to see how to set this up.) The first two litters above look like they might fit this kind of cross.
If you do an aabb x Aabb cross, or an aabb x aaBb cross, you should get 1/2 affected puppies. The third litter above looks like this.
If you do an aabb x AABB, or AaBB, or AABb cross, you shouldn't get any affected puppies, even though two of these possible normal dogs are carrying one deleterious allele each.
If you manage to set up an AaBb x AaBb breeding, you should get a 15 normal : 1 affected ratio, and that will cinch the deal. Figuring out which animals are probably AaBb carriers is left as an exercise for the breeder . . . Seriously, you can guess at this by breeding affected animals to potential carriers and looking for a 1 : 4 ratio in the puppies, since the normal animals involved in these breedings are probably AaBb carriers, rather than one of the other possible kinds of carriers.
At this point, however, you're getting into a mess and may want to switch back to pedigree analysis and see what you can figure out from animals that were bred in the past. An affected x carrier breeding that took place thirty years ago is as good as one you do yourself -- if you can trust the records and pedigrees you have on that breeding and the resultant litter.
Extra complications:
a) Suppose you see a markedly different presentation with different animals: for example, differences in age of onset or severity or exact types of symptoms. This might suggest a trinucleotide-repeat disorder, or variable expressivity due to varying environmental conditions or modifier genes, or the simultaneous existence of multiple independent syndromes.
The same clinical form of human diabetes can be caused by 15 different mutations on chromosome 7 or by mutations on chromosome 20 (Isabell, An Introduction to Genetics for Dog Breeders). These forms of diabetes would be inherited independently, even if one family was unlucky enough to suffer from more than one. Epilepsy which affects predominantly males, shows onset at about age 2-3 years, and causes infrequent but severe grand mal seizures is not likely to be genetically related to epilepsy which affects both sexes equally, shows onset at about age 12-18 months, and causes fly-biting syndrome.
b) Conversely, if a fair number of animals are affected by something, but not by the same thing, consider the underlying metabolic pathways, if possible. Is it possible that the seizures, skin disorders, and an obsessive-compulsive behavior disorder that you see in a breed might all be caused by the same underlying problem with copper metabolism? (Dodman suggests such a system in bull terriers). Hydrocephalus (at least some forms) and syringomyelia are by some suspected to be, possibly, different reflections of the same genetic disorder. Or if your breed suffers from several different immune system dysfunctions, is it possible that some of the different forms of immune disorder all trace back to one underlying genetic problem?
If a particular genetic disorder causes wildly-variant conditions, you may still suspect their underlying genetic connection if they all occur in the same families or lines, rather than one in one line and another in a different line. If it's possible to trace a particular metabolic pathway that, if disabled, could result in all the conditions that you suspect, then their underlying genetic identity becomes more strongly suggested. It may take expert advice from a veterinarian or your friendly neighborhood physiologist to assess this possibility. If you treat all possibly-related diseases as identical in a pedigree, do you get a Mendelian pattern?
Whatever pattern you eventually decide you are seeing, how do you then go on to deal with the trait in your actual real-world breeding program?